| iPsychology
|
|
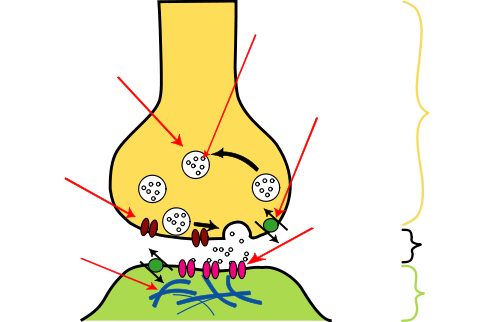
Synapse Synapses are specialized inter neuronal contacts which can be electrical or chemical. Electrical synapses function by the propagation of electrical impulses and do not require elaborate structural organization of the plasma membrane and cytoplasmic organelles. These synapses, which are rare in mammals but common in lower vertebrates, are formed by the close apposition of the cell membranes at a gap junction. Chemical synapses, in contrast, utilize various substances neuro transmitters and neuro peptides for intercellular communication: this chemical transmission requires a sophisticated subcellular mechanism well demonstrated by electron microscopy. By light microscopy, using silver impregnation techniques, only the profile of the end-bulb or button terminal of the axon abutting onto the surface of the perikaryon or dendrite can be seen. A synapse is composed of three constituents: the presynaptic element, the synaptic cleft and the postsynaptic component. Both the presynaptic and the postsynaptic membranes display densities along the specified stretch of cell membrane and these, together with the intervening cleft, are referred to as the synaptic junction. Originally two types of synapse were distinguished on ultra structural examination of the pyramidal cells of the cerebral cortex. The type I junction is usually extensive, the postsynaptic density prominent, rendering the synapse asymmetrical, and the wide synaptic cleft contains a dense plaque. In the type Ii synapse, in contrast, the postsynaptic density is less readily identifiable and the narrower cleft does not contain a dense plaque; the overall configuration is symmetrical. The presynaptic element in most cases is an axon, either at its end ( button terminal ) or along its course ( bout on en passant ): the axonal terminal can form a synapse with any part of another neuron and thus axo-dendritic, axo-somatic and axoaxonal synapses are distinguished. The presynaptic element, however, does not need to be an axonal terminal and synapses can be formed between any part of two neurons, providing a wide variety of synapses. The presynaptic element contains the synaptic vesicles which, in turn, are filled with neurotransmitters. The size, shape and content of the vesicles vary, depending upon the type of synapse. The most frequently Occurring vesicles are apparently clear, spherical and 40-50 nm in diameter. Elongated or flattened vesicles with clear centers are 20 nm wide and 50 nm long. There have been many attempts to correlate vesicular shape with functional activity: spherical vesicles occur in excitatory, type I synapses, whilst flattened vesicles are associated with inhibitory, type II synapses. Larger vesicles of up to 60nm in diameter with a dense core are encountered in areas of catecholamine activity and contain noradrenaline, dopamine or 5hydroxytryptamine. Another, larger dense-cored vesicle, up to lO nm with a spherical dense core of 50-70 nm in diameter, is most frequently seen in the presynaptic terminals of autonomic ganglia, but also occurs in various parts of the central nervous system intermingled with clear vesicles. The presynaptic element, particularly the axonal terminal, also contains other organelles, including mitochondria, neurofilaments, microtubules, smooth endoplasmic reticulum and glycogen. The synaptic cleft, separating the presynaptic and postsynaptic membranes, is 20-30 nM wide and usually contains a dense plaque of intercellular material. In addition, filaments appear to traverse the cleft and contribute to the formation of the plaque. The postsynaptic membrane is rendered conspicuous by an accumulation of dense material on its cytoplasmic surface. This post-synaptic density, composed of granular material and an occasional filamentous structure, is more pronounced in asymmetrical, type I, synapses. The existence of postsynaptic density is dependent upon the presence of the presynaptic element: if the presynaptic structures degenerate, the postsynaptic density will eventually disappear. The postsynaptic element so contains cisternac of the smooth endoplasmic reticulum and the spine apparatus, which is composed of 2-3 flattened cisternac, separated by plaques of dense material. The morphological appearances of synapses reflect the functional dynamics of neurotransmission. Accordingly, in chemical synapses the vesicles contain quanta of the neurotransmitter which is discharged into the synaptic cleft after the vesicle has fused with the presynaptic membrane. Thus, the uptake of calcium into the nerve terminal triggers off a chain of events leading to exocytosis: vesicular apposition, membrane fusion and fission. Although this vesicular or exocytotic hypothesis of neurotransmission provides a plausible explanation for the quintal nature of transmitter release, an increasing body of evidence now suggests that this not be the only mechanism by which chemical signals in the central nervous system are propagated. Chemical substances may be released from non-synaptic axon terminals and even from regions of nerve cells other than presynaptic axon. A parasynaptic system is envisaged in which so called neuroactive or informational substances reach specific target cell receptors by diffusion from release points through the extracellular space. The mechanism of exocytosis itself remains poorly understood not only morphologically, but also neurochemistry, and various alternative biochemical mechanisms have been considered to explain the release of neurotransmitters. These are contact points, usually between axons and dendrites, perikaryons, or rarely, other axons. The axon terminates in a rounded structure called a bouton terminaux. The actual synapse is formed by apparently thickened presynaptic and postsynaptic membranes with a 20-30 nM cleft between. The thickening is actually due to a dense layer of cytoplasmic proteins and filaments underlying the synaptic membranes. Each bouton contains numerous vesicles containing neurotransmitters which are released with the arrival of an action potential, then diffuse across the synapse and act upon receptors on the postsynaptic membrane. Subsequently, this membrane depolarizes and another action potential is generated.
|
|
